Хромозоми
Дефиниција - шта су хромозоми?
Генетска грађа ћелије чува се у облику ДНК (деоксирибонуклеинска киселина) и њених база (аденин, тимин, гванин и цитозин). У свим еукариотским ћелијама (животиња, биљака, гљивица) ово је присутно у ћелијском језгру у облику хромозома. Хромозом се састоји од једног, кохерентног молекула ДНК, који је повезан са одређеним протеинима.
Назив хромозом је изведен из грчког и може се приближно превести као „тело у боји“. Ово име потиче из чињенице да су врло рано у историји цитологије (1888) научници успели да их обоје помоћу посебних основних боја и идентификују у светлосном микроскопу. Међутим, они су заиста видљиви само у одређеној тачки ћелијског циклуса, митози (у заметним ћелијама, мејози), када је хромозом посебно густ (згуснут).

Како се граде хромозоми?
Ако би цела ДНК двострука завојница ћелије, тј. Око 3,4 к 109 базних парова, била повезана заједно, то би резултирало дужином од преко једног метра. Укупна дужина свих додатих хромозома је само око 115 µм. Ова разлика у дужини се објашњава врло компактном структуром хромозома, у којима се ДНК неколико пута намотава или спиралира на врло специфичан начин.
Хистони, посебан облик протеина, играју важну улогу у томе. Укупно постоји 5 различитих хистона: Х1, Х2А, Х2Б, Х3 и Х4. Два од последња четири хистона комбинују се и формирају цилиндричну структуру, октамер, око које се двострука завојница навија два пута (= супер завојница). Х1 се везује за ову структуру како би је стабилизовао.
Овај комплекс ДНК, октамера и Х1 назива се нуклеозом. Неколико ових нуклеозома је сада „попут низа бисера“ у релативно кратким интервалима (10-60 базних парова) један иза другог. Делови између хромозома називају се одстојном ДНК. Појединачни нуклеозоми сада поново долазе у контакт преко Х1, што ствара даље спиралирање, а тиме и компресију.
Добијени ланац је заузврат у петљама које су стабилизоване окосницом направљеном од киселих нехистонских протеина, такође познатих као хертони. Ове петље су заузврат присутне у спиралама стабилизованим протеинима, што резултира последњом фазом компресије. Међутим, овај високи степен компресије јавља се само у контексту деобе ћелија током митозе.
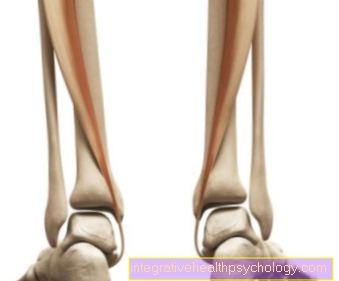
У овој фази можете видети и карактеристичан облик хромозома, који се састоји од две хроматиде. Место њиховог повезивања назива се центромера. Подељује сваки метафазни хромозом на два кратка и два дугачка крака, која се називају и п и к кракови.
Ако центромера лежи приближно у средини хромозома, она се назива метацентрични хромозом, ако лежи у потпуности на једном од крајева акроцентричног хромозома. Они између се називају субметацентрични хромозоми. Ове разлике, које се већ могу видети под светлосним микроскопом, заједно са дужином, омогућавају почетну класификацију хромозома.
Који су теломери?
Теломери су крајеви хромозома са понављајућим секвенцама (ТТАГГГ). Они не садрже никакве релевантне информације, већ служе за спречавање губитка релевантнијих ДНК делова. Са сваком поделом ћелије, део хромозома се губи механизмом репликације ДНК.
Дакле, теломери су у извесном смислу пуфер који одлаже тачку у којој ћелија дељењем губи важне информације. Ако су теломери ћелије дужи од 4.000 базних парова, покреће се програмирана ћелијска смрт (апоптоза). Ово спречава ширење неисправног генетског материјала у организму. Неколико ћелија има теломеразе, тј. Ензиме који су способни да поново продуже теломере.
Поред матичних ћелија, из којих произилазе све остале ћелије, то су полне ћелије и одређене ћелије имуног система. Поред тога, теломеразе се такође налазе у ћелијама карцинома, због чега се у овом контексту говори о имортализацији ћелије.
Све о теми прочитајте овде: Теломери - Анатомија, функција и болести
Шта је хроматин?
Хроматин се односи на целокупан садржај ћелијског језгра који може бити обојен базом. Стога, поред ДНК, термин укључује и одређене протеине, нпр. Хистоне и хертоне (види структуру), као и одређене фрагменте РНК (хн и снРНА).
Зависно од фазе у ћелијском циклусу или од генетске активности, овај материјал је доступан у различитим густинама. Густи облик се назива хетерохроматин. Да би га било лакше разумети, могао би се стога сматрати „обрасцем за складиштење“ и овде опет направити разлику између конститутивног и факултативног хетерохроматина.
Конститутивни хетерохроматин је најгушћи облик, који је присутан у свим фазама ћелијског циклуса у свом највишем степену кондензације. Чини око 6,5% људског генома и углавном се налази у близини центромера и крајева кракова хромозома (теломера), али и на другим местима (углавном хромозомима 1, 9, 16, 19 и И) . Поред тога, већина конститутивног хетерохроматина налази се у близини нуклеарне мембране, односно на ивицама ћелијског језгра. Простор у средини резервисан је за активни хроматин, еухроматин.
Факултативни хетерохроматин је мало мање густ и може се активирати и деактивирати по потреби или у зависности од фазе развоја. Добар пример за то је други Х хромозом у женским кариотипима. С обзиром да је један Кс хромозом у основи довољан да ћелија преживи, као што је на крају довољан и за мушкарце, један од два се деактивира у ембрионалној фази. деактивирани Кс хромозом познат је као Баррово тело.
Тек током деобе ћелија, у контексту митозе, она се потпуно кондензује, при чему достиже највећу компресију у метафази. Међутим, с обзиром да се различити гени често различито читају - уосталом, нису сви протеини увек потребни у истој количини - овде се такође прави разлика између активног и неактивног еухроматина.
Прочитајте више о овоме испод: Хроматин
Хаплоидни хромозоми
Хаплоид (грчки хаплоос = појединачно) значи да су сви хромозоми ћелије присутни појединачно, тј. Не у паровима (диплоидни) као што је то обично случај. Ово је природно стање свих ћелија јајних ћелија и сперме, у којима се две идентичне хроматиде у почетку не раздвајају током прве мејозе, већ се прво раздвајају сви парови хромозома.
Као резултат, после прве мејозе, ћерке ћелије код људи имају само 23 уместо уобичајених 46 хромозома, што одговара половини хаплоидног сета хромозома. Пошто ове ћерке ћелије још увек имају идентичну копију сваког хромозома који се састоји од 2 хромозома, потребна је друга мејоза у којој су две хроматиде одвојене једна од друге.
Политенски хромозоми
Политенски хромозом је хромозом који се састоји од великог броја генетски идентичних хроматида. Пошто су такви хромозоми лако уочљиви и под мањим увећањем, понекад се називају џиновским хромозомима. Предуслов за ово је ендорепликација, у којој се хромозоми унутар ћелијског језгра множе неколико пута, а да се дељење ћелија не догоди.
Које су функције хромозома?
Хромозом као организациона јединица нашег генома првенствено служи да осигура да се удвостручени геном равномерно распореди међу ћерке ћелије током деобе ћелија. Да бисте то урадили, вреди пажљивије погледати механизме ћелијске деобе или ћелијског циклуса:
Ћелија проводи већи део ћелијског циклуса у интерфази, што значи читав временски период у којем ћелија неће одмах да се подели. Ово је заузврат подељено на фазе Г1, С и Г2.
Г1 фаза (Г као у празнини, тј. Јаз) одмах следи поделу ћелије. Овде се ћелија поново повећава у величини и врши опште метаболичке функције.
Одавде се такође може пребацити у Г0 фазу. То значи да се она мења у стадијум који више није способан да се дели и, у нормалним случајевима, такође се увелико мења како би се испунила врло специфична функција (диференцијација ћелија). Да би се испунили ови задаци, врло специфични гени се читају интензивније, други мање или никако.
Ако сегмент ДНК није потребан дуго времена, често се налази у деловима хромозома који су дуго били густо упаковани (види хроматин). С једне стране, ово има за циљ уштеду простора, али поред осталих механизама регулације гена, то је и додатна заштита од случајног читања. Међутим, такође је примећено да под врло специфичним условима диференциране ћелије из Г0 фазе могу поново да уђу у циклус.
Након Г1 фазе следи С фаза, односно фаза у којој се синтетише нова ДНК (репликација ДНК). Овде целокупна ДНК мора бити у свом најлабавијем облику, тј. Сви хромозоми су потпуно одмотани (види структуру).
На крају фазе синтезе, цео генетски материјал је у дупликату присутан у ћелији. Будући да је копија још увек причвршћена за оригинални хромозом преко центромере (види структуру), не говори се о дуплицирању хромозома.
Сваки хромозом се сада састоји од две хроматиде уместо од једне, тако да касније може да поприми карактеристични облик Кс током митозе (строго говорећи, облик Кс односи се само на метацентричне хромозоме). У наредној Г2 фази одвија се непосредна припрема за поделу ћелија. То такође укључује детаљну проверу грешака у репликацији и прелома жица, које се могу поправити ако је потребно.
У основи постоје две врсте деобе ћелија: митоза и мејоза. Са изузетком полних ћелија, све ћелије организма настају митозом, чији је једини задатак формирање две генетски идентичне ћерке ћелије.
Мејоза, с друге стране, има за циљ генерирање генетски различитих ћелија:
У првом кораку се поделе одговарајући (хомологни), али не и идентични хромозоми. Тек у следећем кораку хромозоми, који се састоје од две идентичне хроматиде, одвајају се и поново дистрибуирају по две ћерке ћелије, тако да на крају из једне ћелије претече настају четири полне ћелије са различитим генетским материјалом.
Облик и структура хромозома су од суштинске важности за оба механизма: Посебне „протеинске нити“, такозвани вретенски апарат, прикаче се на високо кондензоване хромозоме и извлаче хромозоме у фино регулисаном процесу из средње равни (екваторијална раван) на супротне полове ћелије око једног како би се обезбедила равномерна расподела. Чак и мале промене у микроструктури хромозома овде могу имати озбиљне последице.
У свих сисара, однос полних хромозома Кс и И такође одређује пол потомака. У основи, све зависи од тога да ли сперматозоиди који се сједине са јајном ћелијом имају Кс или И хромозом. Пошто се оба облика сперме увек производе у потпуно истој мери, вероватноћа је увек уравнотежена за оба пола. Овај случајни систем гарантује равномернију расподелу полова него што би то био случај, на пример, са факторима околине као што је температура.
Сазнајте више о теми: Подела ћелијског језгра
Како се генетски састав преноси путем хромозома?
Данас знамо да се особине наслеђују преко гена који се чувају у ћелијама у облику ДНК. Они су заузврат подељени у 46 хромозома, на којима је распоређено 25 000-30000 људских гена.
Поред самог својства, које се назива фенотип, постоји и генетски еквивалент, који се назива генотип. Место где се ген налази на хромозому назива се локус. Пошто људи имају двоструко сваки хромозом, сваки ген се такође јавља два пута. Једини изузетак од тога су Кс-хромозомски гени код мушкараца, јер И-хромозом носи само делић генетских информација пронађених на Кс-хромозому.
Различити гени који се налазе на истом локусу називају се алелима. Често постоји више од два различита алела у једном локусу. Тада се говори о полиморфизму. Такав алел може једноставно бити безопасна варијанта (нормална варијанта), али и патолошке мутације које могу бити покретач наследне болести.
Ако је мутација једног гена довољна за промену фенотипа, говори се о моногенском или менделовском наслеђивању. Међутим, многе наследне особине се наслеђују кроз неколико гена у интеракцији и стога их је много теже проучити.
Будући да мајка и отац сваки од својих два гена преносе детету у менделско наследство, у следећој генерацији увек постоје четири могуће комбинације, при чему оне такође могу бити исте у односу на једно својство. Ако оба алела појединца имају исти ефекат на фенотип, појединац је хомозиготан у односу на ову карактеристику и карактеристика је у потпуности изражена.
Хетерозиготи имају два различита алела који могу међусобно да комуницирају на различите начине: Ако је један алел доминантан над другим, у потпуности потискује свој израз и доминантна особина постаје видљива у фенотипу. Потиснути алел назива се рецесивним.
У случају кодоминантног наслеђа, оба алела се могу изразити међусобно непромењена, док у случају средњег наследства постоји мешавина обе карактеристике. Добар пример за то је систем крвних група АБ0, у којем су А и Б међусобно доминантни, али 0 доминирају један над другим.
Који је нормалан скуп хромозома код људи?
Људске ћелије имају 22 полно независна пара хромозома (аутосоми) и два полна хромозома (гоносоми), па укупно 46 хромозома чини један сет хромозома.
Аутосоми обично долазе у паровима. Хромозоми пара су слични по облику и секвенци гена и због тога се називају хомологним. Два Кс хромозома код жена су такође хомологна, док мушкарци имају Кс и И хромозом. Они се разликују у облику и броју присутних гена на такав начин да се више не може говорити о хомологији.
Герм ћелије, тј. Јајне ћелије и сперматозоиди, имају само половину комплета хромозома услед мејозе, наиме по 22 појединачна аутосома и по један гоносом. Пошто се полне ћелије стапају током оплодње и понекад замењују читаве сегменте (укрштање), ствара се нова комбинација хромозома (рекомбинација). Сви хромозоми заједно називају се кариотип, који је са неколико изузетака (види аберације хромозома) идентичан код свих јединки истог пола.
Овде можете сазнати све о тој теми: Митоза - једноставно објашњено!
Зашто увек постоје парови хромозома?
У основи, на ово питање се може одговорити једном реченицом: Зато што се показало корисним.Присуство парова хромозома и принцип рекомбинације су од суштинског значаја за наслеђивање у погледу полне репродукције. На овај начин, потпуно нова јединка може случајно настати из генетског материјала две јединке.
Овај систем енормно повећава разноликост својстава унутар врсте и осигурава да се може много брже и флексибилније прилагодити промењеним условима околине него што би то било могуће само мутацијом и селекцијом.
Двоструки сет хромозома такође има заштитни ефекат: ако би мутација гена довела до отказивања функције, у другом хромозому још увек постоји нека врста „резервне копије“. То није увек довољно да организам надокнади квар, посебно ако је мутирани алел доминантан, али повећава шансу за то. Поред тога, на овај начин се мутација не преноси аутоматски на све потомке, што заузврат штити врсту од претерано радикалних мутација.
Шта је мутација хромозома?
Генетски дефекти могу настати због јонизујућег зрачења (нпр. Рендгенски зраци), хемијских супстанци (нпр. Бензопирен у цигаретном диму), одређених вируса (нпр. ХП вируса) или, са малом вероватноћом, могу настати и сасвим случајно. У његовом развоју често учествује неколико фактора. У принципу, такве промене могу да се појаве у свим телесним ткивима, али из практичних разлога анализа је обично ограничена на лимфоците (посебна врста имуних ћелија), фибробласте (ћелије везивног ткива) и ћелије коштане сржи.
Мутација хромозома је главна структурна промена у појединачним хромозомима. С друге стране, одсуство или додавање целих хромозома представљало би мутацију генома или плоидије, док се термин мутација гена односи на сразмерно мале промене унутар гена. Појам аберација хромозома (латински аберраре = одступати) је нешто шири и укључује све промене које се могу детектовати светлосним микроскопом.
Мутације могу имати врло различите ефекте:
- Тихе мутације, тј. Мутације у којима промена нема ефекта на појединца или њихово потомство, прилично су нетипичне за хромозомске аберације и чешће се налазе у подручју мутација гена или тачке.
- Говори се о мутацији губитка функције када мутација резултира погрешно склопљеним и самим тим нефункционалним протеином или уопште без њега.
- Такозване мутације добитка функције мењају врсту ефекта или количину произведених протеина на такав начин да настају потпуно нови ефекти. С једне стране, ово је пресудан механизам за еволуцију, а тиме и за опстанак врсте или појаву нових врста, али с друге стране, као у случају хромозома из Филаделфије, такође може пресудно допринети развој ћелија карцинома.
Најпознатији од различитих облика аберација хромозома су вероватно нумеричке аберације, у којима су појединачни хромозоми присутни само једном (моносомија) или чак троструко (трисомија).
Ако се ово односи само на један хромозом, он се назива анеуплоидија, а на читав низ хромозома утиче полиплоидија (три- и тетраплоидија). У већини случајева, ова неправилна расподела настаје током развоја полних ћелија нераздвајањем (нераздвајањем) хромозома током ћелијске деобе (мејоза). То доводи до неравномерне расподеле хромозома на ћеркама и тиме до нумеричке аберације код детета.
Моносомије несполних хромозома (= аутосоми) су некомпатибилне са животом и зато се не јављају код живе деце. Изузев трисомија 13, 18 и 21, аутосомне трисомије готово увек доводе до спонтаних побачаја.
У сваком случају, за разлику од аберација полних хромозома, које такође могу бити неупадљиве, увек постоје озбиљни клинички симптоми и, по правилу, мање или више изражене спољне абнормалности (дисморфизми).
Таква неправилна расподела може се десити и касније у животу са поделом митотичких ћелија (све ћелије осим полних ћелија). Пошто поред погођених ћелија постоје и непромењене ћелије, говори се о соматском мозаику. Под соматским (грчки сома = тело) подразумевају се све ћелије које нису заметне ћелије. Пошто је захваћен само мали део ћелија тела, симптоми су обично много блажи. Стога типови мозаика често остају неоткривени дуго времена.
Овде можете сазнати све о тој теми: Хромозомска мутација
Шта је хромозомска аберација?
Структурна аберација хромозома у основи одговара дефиницији мутације хромозома (види горе). Ако количина генетског материјала остане иста и једноставно се другачије дистрибуира, говори се о уравнотеженој аберацији.
То се често врши транслокацијом, тј. Преносом сегмента хромозома у други хромозом. Ако је реч о размени између два хромозома, један говори о реципрочној транслокацији. Будући да је за производњу протеина потребно само око 2% генома, вероватноћа је да је такав ген на тачки прелома и тиме губи своју функцију или је у њему оштећена. Стога таква уравнотежена аберација често пролази непримећено и преноси се кроз неколико генерација.
Међутим, то може довести до неправилне расподеле хромозома током развоја полних ћелија, што може довести до неплодности, спонтаних побачаја или потомства са неуравнотеженом аберацијом.
Неуравнотежена аберација такође може да се догоди спонтано, тј. Без породичне историје. Вероватноћа да ће се дете родити живо са неуравнотеженом аберацијом у великој мери зависи од хромозома који су погођени и варира између 0 и 60%. То доводи до губитка (= брисања) или дуплирања (= дуплирања) сегмента хромозома. У овом контексту, такође се говори о делимичним моно- и трисомијама.
У неким случајевима се јављају заједно у два различита региона, при чему је делимична моносомија обично пресуднија за појаву клиничких симптома. Ово су истакнути примери брисања Синдром мачјег крика и синдром Волф-Хирсцххорн.
Говори се о микроделецији када се промена више не може утврдити светлосним микроскопом, тј. Када је реч о губитку једног или неколико гена. Сматра се да је овај феномен узрок Прадер-Виллијевог синдрома и Ангелмановог синдрома и уско је повезан са развојем ретионобластома.
Робертсонова транслокација је посебан случај:
Два акроцентрична хромозома (13, 14, 15, 21, 22) се уједињују у својој центромери и формирају један хромозом након што се изгубе кратки кракови (види структуру). Иако ово резултира смањеним бројем хромозома, ово се назива уравнотеженом аберацијом, јер се губитак кратких кракова ових хромозома може лако надокнадити. И овде су ефекти често приметни само у следећим генерацијама, јер је врло велика вероватноћа побачаја или живе деце са трисомијом.
Ако постоје две паузе унутар хромозома, може се догодити да се међусегмент ротира за 180 ° и угради у хромозом. Овај процес, познат као инверзија, неуравнотежен је само ако се тачка прелома налази унутар активног гена (2% укупног генетског материјала). У зависности од тога да ли је центромера унутар или изван обрнутог сегмента, то је пери- или парацентрична инверзија. Ове промене такође могу допринети неравномерној расподели генетског материјала на полним ћелијама.
У парацентричној инверзији, у којој центромера није у обрнутом сегменту, могу се појавити и заметне ћелије са две или без центромере. Као резултат, одговарајући хромозом се губи током првих дељења ћелија, што готово сигурно доводи до побачаја.
Уметање је уградња фрагмента хромозома на неко друго место. И овде су на сличан начин првенствено погођени потомци. Прстенасти хромозом се може јавити нарочито након брисања крајњих делова. Тип и величина секвенци су одлучујући за тежину симптома. Поред тога, то може довести до нетачне расподеле, а тиме и до врста мозаика у ћелијама тела.
Ако се метафазни хромозом током раздвајања ћелија нетачно одвоји, могу доћи до изохромозома. То су два потпуно иста хромозома која се састоје од само дугих или само кратких кракова. У случају Кс хромозома, ово се може манифестовати као Улрицх-Турнер синдром (моносомија Кс).
Прочитајте више информација о овој теми: Хромозомска аберација
Трисомија 21
Трисомија 21, познатија као Даунов синдром, вероватно је најчешћа нумеричка хромозомска аберација међу живорођенима, с тим што су мушкарци погођени нешто чешће (1,3: 1).
Вероватноћа настанка трисомије 21 зависи од различитих демографских фактора, као што је просечна старост рођења мајки, и незнатно варира од региона до региона.
95% трисомије 21 настаје као резултат грешке у подели у контексту мејозе (деоба полних ћелија), наиме недисјункције, тј. Неуспеха у одвајању сестринских хроматида.
Они су познати као слободне трисомије и настају 90% у мајке, 5% у очини и још 5% у ембрионалном геному.
Још 3% је резултат неуравнотежене транслокације било на хромозому 14 или као 21; 21 транслокацију, стварајући нормалан и двоструки хромозом 21. Преосталих 2% су врсте мозаика код којих трисомија није настала у полним ћелијама и стога не утиче на све телесне ћелије. Типови мозаика су често толико благи да могу дуго остати потпуно неоткривени.
У сваком случају, треба извршити хромозомско испитивање како би се разликовала симптоматски идентична слободна трисомија од могуће наслеђене транслокационе трисомије. Тада може да следи породична историја претходних генерација.
Да ли вас занима ова тема? Прочитајте следећи чланак о овоме: Трисомија 21
Трисомија 13
Трисомија 13 или Патауов синдром има фреквенцију 1: 5000 и много је ређи од Довновог синдрома. Узроци (слободне трисомије, транслокације и врсте мозаика) и њихова процентуална дистрибуција су углавном идентични.
У теорији, скоро сви случајеви могу се дијагностицирати пренатално помоћу ултразвука или ПАПП-А теста. Будући да тест ПАПП-А није нужно део рутинских прегледа, око 80% случајева у Централној Европи дијагностикује се пре рођења.
На ултразвуку се већ могу видети остаци израслине, обострана пукотина усне и непца и необично мале очи (микрофталмија). Поред тога, обично су присутне малформације предњег мозга и лица различитог степена озбиљности (холопросенцефалија).
Док су у лобарном облику мождане хемисфере готово потпуно одвојене и стварају се бочне коморе, у полулобарном облику често је одвојен само задњи део мозга и недостају бочне коморе. У најтежем облику, алобарном облику, нема раздвајања можданих хемисфера.
Дојенчад са полу- или алобарним обликом обично умиру одмах након рођења. После месец дана, стопа смртности је око 50% живорођених. До 5. године живота стопа морталитета од тризомије 13 повећава се на 90%. Због малформација у мозгу, у већини случајева болесни људи остају везани за кревет цео живот и не могу да говоре, због чега су зависни од пуне неге. Поред тога, могу постојати и далекосежне физичке манифестације Трисмоие 13.
Прочитајте више о томе на: Трисомија 13 код нерођеног детета
Трисомија 16
У основи, трисомија 16 је најчешћа трисомија (око 32% свих трисомија), али жива деца са трисомијом 16 су врло ретка. Генерално, живорођавање се дешава само у делимичним врстама трисомија или мозаика. Међутим, међу трисомијама је најчешће одговорна за мртворођене деце: 32 од 100 побачаја због хромозомских аберација могу се пратити до овог облика трисомије.
Стога су документоване углавном пренаталне, тј. Пренаталне карактеристике које се могу идентификовати. Овде се истичу разне срчане мане, успорени раст, појединачна пупчана артерија (иначе двострука) и повећана прозирност врата, што се објашњава накупљањем течности услед још увек не потпуно развијеног лимфног система и повећане еластичности коже на овом подручју. Поред тога, физиолошка пупчана кила, тј. Привремено померање великог дела црева кроз пупак према споља, често се не регресира правилно, што је познато као пуцање омфалокеле или пупчане врпце.
Флектурна контрактура прекрижених прстију такође се често може открити на ултразвуку. Код неколико живорођених, примећује се генерализована хипотензија мишића, тј. Општа слабост мишића. То доводи до слабости пијења и може осигурати вештачко храњење дојенчета. Често се јавља и бразда са четири прста која је тако карактеристична за трисомије. И овде је учесталост појаве трисомије у директној вези са узрастом мајке.
Трисомија 18
Едвардсов синдром, тј. Трисомија 18, јавља се са фреквенцијом 1: 3000. Са пренаталном дијагностиком исто је као и са Патауовим синдромом: и овде би исти прегледи омогућили да се сви пацијенти у потпуности пронађу пре рођења. Узроци и њихова дистрибуција могу се упоредити са другим трисомијама (видети трисомију 21).
Поред тога, делимичне трисомије се јављају у трисомији 18, које, попут типова мозаика, доводе до много блажих клиничких токова. Повезани дисморфизми су такође изузетно карактеристични за Едвардсов синдром: При рођењу пацијенти имају знатно смањену телесну тежину од 2 кг (нормално: 2,8-4,2 кг), широко чело које се повлачи, генерално неразвијену доњу половину лица са малим устима отвор, уски прорези капака и окренути уназад, уши промењене у облику (фауново ухо). Приметан је и задњи део главе, који је необично јако развијен за новорођенче. Ребра су необично уска и крхка. Новорођенчад такође има трајну напетост (тонус) целе мускулатуре, која, међутим, код преживелих назадује након првих неколико недеља.
Још једна карактеристична карактеристика је прелазак 2. и 5. прста преко 3. и 4. са укупним бројем закуцаних прстију, док су стопала необично дугачка (протегнута), имају посебно изражену пету, закржљали нокти на ногама и уназад постављени ножни палац .
Озбиљне малформације органа су честе и обично се јављају у комбинацији: оштећења срца и бубрега, неправилно савијање (малротација) црева, адхезије перитонеума (месентериум цоммуне), зачепљење једњака (атрезија једњака) и још много тога.
Због ових малформација, стопа смртности је око 50% у прва 4 дана, само око 5-10% живи до преко годину дана. Преживљавање у одраслој доби апсолутни је изузетак, у сваком случају, интелектуални инвалидитет је веома изражен и не може да говори, прикован је за кревет и инконтинентан, па у потпуности зависи од спољне помоћи.
За детаљније информације о трисомији 18, такође прочитајте наш детаљни чланак на ту тему:
- Трисомија 18 (Едвардсов синдром)
- Трисомија 18 код нерођеног детета
Трисомија Кс
Трисомија Кс је најнеупадљивији облик нумеричке хромозомске аберације, изглед оболелих, који су логично све жене, не разликује се много од осталих жена. Неки се истичу зато што су посебно високи и имају донекле „пунашне“ црте лица. Ментални развој такође може бити углавном нормалан, у распону од граничног нормалног до благог менталног инвалидитета.
Међутим, овај дефицит интелигенције је нешто озбиљнији него код осталих трисомија полних хромозома (КСКСИ и КСИИ). Са фреквенцијом од 1: 1000 то заправо није тако ретко, али пошто трисомија обично није повезана са клинички значајним симптомима, већини жена са болешћу вероватно никада неће бити постављена дијагноза током целог живота.
Носачи се углавном откривају случајно током породичног прегледа или током пренаталне дијагностике.Плодност се може мало смањити, а стопа аберација полних хромозома у следећој генерацији може се мало повећати, тако да се препоручује генетско саветовање ако желите да имате децу.
Као и код осталих трисомија, трисомија Кс се најчешће развија као слободна трисомија, тј. Због недостатка поделе (недисјункције) сестринских хроматида. И овде се обично јавља током сазревања јајних ћелија мајке, мада се вероватноћа повећава са годинама.
Крхки Кс синдром
Крхки Кс синдром или Мартин Беллов синдром је пожељнији код мушкараца, јер имају само један Кс хромозом и због тога су више погођени променом.
Јавља се са фреквенцијом од 1: 1250 међу живорођеним мушким породима у једној години, што га чини најчешћим обликом неспецифичне менталне ретардације, тј. Свим менталним хендикепима који се не могу описати посебним синдромом са типичним знацима.
Крхки Кс синдром се обично може јавити код девојчица у нешто слабијем облику, што је због случајне инактивације једног од Кс хромозома. Што је већи проценат искљученог здравог Кс хромозома, симптоми су јачи.
Међутим, најчешће су жене носиоци премутације, која још увек не производи никакве клиничке симптоме, али масовно повећава вероватноћу потпуне мутације код њихових синова. У врло ретким случајевима мушкарци такође могу бити носиоци премутације, коју потом могу пренети само на ћерке које су, међутим, обично клинички здраве (Схерманов парадокс).
Синдром је покренут изузетно повећаним бројем ЦГГ тројки (одређена основна секвенца) у гену ФМР (крхко место менталне ретардације); уместо 10-50 копија, премутација 50-200, када је потпуно развијена 200- 2000 примерака.
Под светлосним микроскопом ово изгледа као прелом дуге руке, по чему је синдром и добио име. То доводи до деактивирања погођеног гена, што заузврат узрокује симптоме.
Погођене особе показују успорен развој говора и покрета и могу показати проблеме у понашању који могу ићи у правцу хиперактивности, али и аутизма. Чисто спољне абнормалности (знаци дисморфизма) су дуго лице са истакнутом брадом и избоченим ушима. Са пубертетом су тестиси често увелико увећани (макроорхидија), а црте лица постају грубље. Постоји мало нагомилавање психолошких абнормалности и нарочито рана менопауза међу носиоцима премутације.
Шта је анализа хромозома?
Анализа хромозома је процес у цитогенетици помоћу којег се могу открити нумеричке или структурне аберације хромозома.
Таква анализа би се користила, на пример, ако се одмах посумња на хромозомски синдром, односно у случају малформација (дисморфизми) или интелектуалних сметњи (ретардација), али и у случају неплодности, редовног побачаја (побачаја) и такође код одређени карциноми (нпр. лимфоми или леукемија).
За ово су обично потребни лимфоцити, посебна врста имуних ћелија која се добија из крви пацијента. Будући да се на овај начин може добити само релативно мала количина, ћелије се стимулишу да се деле фитохемаглутинином и лимфоцити се затим могу гајити у лабораторији.
У неким случајевима се узимају узорци (биопсије) са коже или кичмене мождине, уз сличан поступак. Циљ је добити што више ДНК материјала који је тренутно у средини ћелијске деобе. У метафази су сви хромозоми распоређени у једном нивоу приближно у средини ћелије, како би се повукли на супротне стране (полове) ћелије у следећем кораку, анафази.
У овом тренутку, хромозоми су посебно тесно упаковани (високо кондензовани). Додан је вретенски отров колхицин, који делује управо у овој фази ћелијског циклуса, тако да се метафазни хромозоми акумулирају. Затим се изолују и обоје помоћу посебних метода бојења.
Најчешћи је ГТГ трак, у коме се хромозоми третирају трипсином, дигестивним ензимом и пигментом Гиемса. Посебно густо набијени региони и они богати аденином и тимином приказани су тамно.
Добијени Г-опсези су карактеристични за сваки хромозом и, поједностављено речено, сматрају се регионима са мање гена. Овако обојена хромозома слика се хиљадуструким увећањем и помоћу рачунарског програма креира кариограм. Поред узорка траке, величина хромозома и положај центромере користе се да би се помогло да се хромозоми распореде у складу са тим. Али постоје и друге методе повезивања које могу имати врло различите предности.
Препоруке уредничког тима
За опште информације погледајте следеће чланке:
- Подела ћелијског језгра
- Функције ћелијског језгра
- Трисомија 21
- Генетске болести

.jpg)